Chapter 6 Autocatalytic Networks
6.1 Why Autocatalytic Networks Matter
One of the central questions in origin-of-life research is how simple chemistry became capable of self-maintenance and evolution.
Many origin-of-life theories focus on replication. In these models, a molecule capable of copying itself is viewed as the starting point of evolution.
However, another possibility exists.
Instead of a single molecule reproducing itself, an entire network of molecules may collectively sustain its own production.
This idea forms the basis of autocatalytic network theory.
In this view:
Life may have begun not with a single replicator, but with a community of mutually supportive molecules.
Autocatalytic networks provide one possible explanation for how organization emerged before the appearance of modern genes.
6.2 What Is Catalysis?
Catalysis occurs when one molecule increases the rate of a chemical reaction without being permanently consumed.
Catalysts are essential in modern biology.
Examples include:
- enzymes accelerating metabolic reactions,
- ribozymes catalyzing RNA reactions,
- mineral surfaces promoting chemical transformations.
Without catalysis, many biological reactions would proceed far too slowly to sustain life.
Catalysis therefore provides a mechanism by which chemical systems can become increasingly organized.
6.3 What Is an Autocatalytic Network?
An autocatalytic network is a collection of molecules in which members of the network help produce one another.
For example:
Molecule A helps produce Molecule B.
Molecule B helps produce Molecule C.
Molecule C helps produce Molecule A.
No single molecule is self-sufficient.
However, the network as a whole becomes self-reinforcing.
This creates an important shift in perspective:
The unit of organization becomes the network rather than the individual molecule.
6.4 Historical Background
The concept of autocatalytic networks became particularly influential through the work of Stuart Kauffman.
Kauffman proposed that sufficiently complex chemical systems may naturally undergo a transition from disconnected reactions to self-sustaining networks.
This idea suggested a possible route to life’s emergence that does not require a highly improbable self-replicating molecule appearing first.
Instead:
- diversity increases,
- catalytic interactions accumulate,
- networks become increasingly connected,
- self-maintaining systems emerge.
This perspective remains an important alternative to purely gene-first theories.
6.5 Network-First Versus Replication-First Models
Two broad approaches to the origin of life can be contrasted.
6.6 Emergence in Chemical Networks
Autocatalytic networks provide an excellent example of emergence.
No individual molecule may possess life-like properties.
However, when many molecules interact:
- feedback loops appear,
- self-maintenance emerges,
- system-level organization develops.
The resulting behavior cannot always be predicted by examining molecules individually.
This illustrates one of the central themes of complexity science:
The whole can exhibit properties that are not obvious from the parts.
6.7 Conceptual Model
In lifesimulatoR, an autocatalytic network is represented using:
- molecular types,
- catalytic relationships,
- abundance changes through time,
- a catalysis matrix.
Although highly simplified, this framework captures the essential idea of mutual reinforcement.
6.8 Creating an Autocatalytic Network
network <- autocatalytic_network(
n_types = 8,
steps = 50,
catalysis_probability = 0.2,
seed = 123
)
names(network)## [1] "time_series" "catalysis_matrix"## # A tibble: 6 × 3
## step molecule abundance
## <int> <chr> <dbl>
## 1 0 M1 0.833
## 2 0 M2 0.504
## 3 0 M3 0.829
## 4 0 M4 0.831
## 5 0 M5 0.815
## 6 0 M6 0.496The output contains both network structure and abundance dynamics.
6.9 Understanding the Catalysis Matrix
The catalysis matrix records which molecules catalyze the production of others.
{r autocatalytic-matrix} id="autocatalytic-matrix" network$catalysis_matrix
The matrix can be viewed as a map of catalytic interactions.
Each connection represents a potential pathway through which molecules influence one another.
6.10 Visualizing Molecular Dynamics
We can examine the abundance of a particular molecular type.
m1 <- subset(
network$time_series,
molecule == "M1"
)
plot_simulation(
m1,
x = "step",
y = "abundance"
)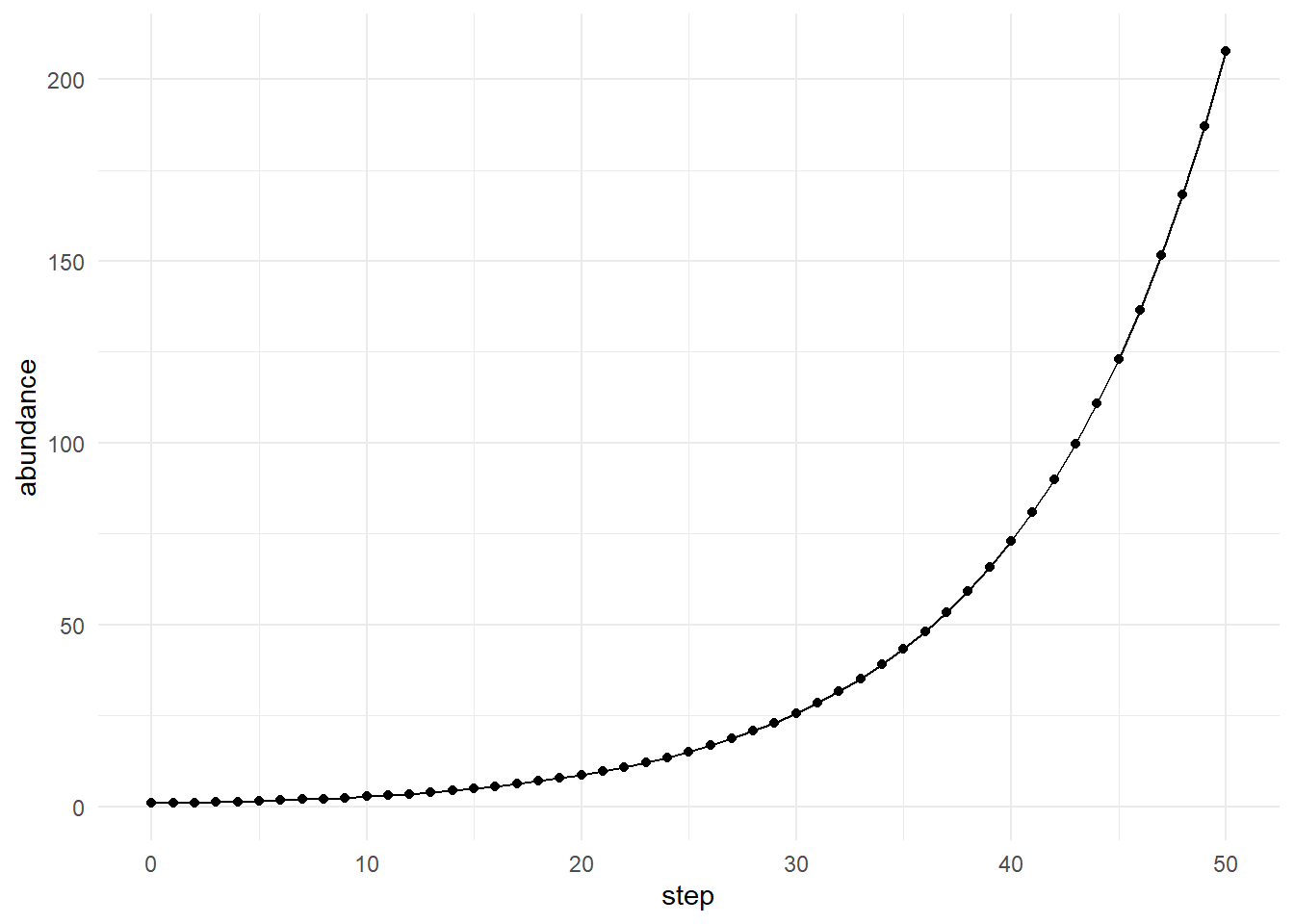
This plot shows how one molecule changes through time as it participates in the broader network.
6.11 Feedback Loops
One of the most important features of autocatalytic systems is the presence of feedback.
6.12 Network Connectivity
An important property of a network is connectivity.
Connectivity refers to how many interactions exist among components.
Low connectivity produces sparse networks.
High connectivity produces dense networks.
The level of connectivity strongly influences network behavior.
6.13 Sparse Versus Dense Networks
sparse_network <- autocatalytic_network(
n_types = 8,
steps = 50,
catalysis_probability = 0.05,
seed = 123
)
dense_network <- autocatalytic_network(
n_types = 8,
steps = 50,
catalysis_probability = 0.4,
seed = 123
)
head(sparse_network$time_series)## # A tibble: 6 × 3
## step molecule abundance
## <int> <chr> <dbl>
## 1 0 M1 0.833
## 2 0 M2 0.504
## 3 0 M3 0.829
## 4 0 M4 0.831
## 5 0 M5 0.815
## 6 0 M6 0.496## # A tibble: 6 × 3
## step molecule abundance
## <int> <chr> <dbl>
## 1 0 M1 0.833
## 2 0 M2 0.504
## 3 0 M3 0.829
## 4 0 M4 0.831
## 5 0 M5 0.815
## 6 0 M6 0.496Dense networks are more likely to contain:
- feedback loops,
- indirect interactions,
- collective dynamics.
Sparse networks tend to be less interconnected.
6.14 The Autocatalytic Threshold
Kauffman proposed that sufficiently connected networks may experience a phase transition.
Below a critical threshold:
- reactions remain largely disconnected.
Above the threshold:
- self-sustaining networks become possible.
This idea is similar to phase transitions observed elsewhere in nature, such as:
- water freezing,
- magnets becoming ordered,
- percolation in physical systems.
The emergence of autocatalytic sets may represent a comparable transition in chemical organization.
6.15 Autocatalytic Networks and Evolution
Autocatalytic networks raise an important question:
Can evolution occur before genetic replication?
Some researchers argue that network-level selection may occur when:
- certain networks persist longer,
- certain networks utilize resources more efficiently,
- certain networks generate more stable structures.
If so, evolution may have begun before modern genes existed.
6.16 Networks Inside Protocells
Autocatalytic networks become even more interesting when combined with compartments.
A protocell can:
- retain catalytic molecules,
- protect network components,
- maintain higher concentrations,
- enable network inheritance during division.
Many modern theories propose that life emerged through interactions among:
- molecular evolution,
- autocatalytic networks,
- protocells.
Rather than one appearing first, these systems may have co-evolved.
6.17 Interpreting Simulation Results
Several outcomes are possible.
6.17.1 Stable Abundances
May indicate:
- balanced network interactions,
- stable catalytic relationships.
6.17.2 Increasing Abundances
May indicate:
- strong positive feedback,
- successful catalytic reinforcement.
6.18 Limitations of the Model
The autocatalytic network model in lifesimulatoR is intentionally simplified.
It does not include:
- realistic reaction kinetics,
- thermodynamic constraints,
- energy requirements,
- environmental variation,
- molecular structure,
- reaction mechanisms,
- resource competition.
Its purpose is to illustrate the principles of network organization and emergence rather than reproduce real chemistry.
6.19 Connections to Other Chapters
This chapter integrates many ideas developed earlier.
Prebiotic chemistry provides molecular diversity.
Molecular evolution introduces information and selection.
Diversity creates opportunities for interaction.
Protocells create compartments.
Autocatalytic networks provide organization.
Together these components illustrate several of the major pathways proposed for the emergence of life.
6.20 Key Takeaways
- Autocatalytic networks consist of mutually reinforcing molecules.
- Organization can emerge from interactions among many components.
- Networks may provide an alternative to purely replication-first origin-of-life theories.
- Feedback loops play a central role in network behavior.
- Connectivity strongly influences network dynamics.
- Self-sustaining systems may emerge above critical network thresholds.
- Autocatalytic networks become especially powerful when combined with protocells.
lifesimulatoRprovides a simplified framework for exploring these ideas.
6.21 Suggested Readings
- Kauffman, S. (1993). The Origins of Order.
- Dyson, F. (1985). Origins of Life.
- Hordijk, W., & Steel, M. (2004). Detecting Autocatalytic Sets.
- Smith, E., & Morowitz, H. (2016). The Origin and Nature of Life on Earth.
- Walker, S. I. (2017). Origins of Life and Complexity.
6.22 Reflection Questions
- How does a network-first model differ from a replication-first model?
- Can organization emerge without genetic information?
- What role do feedback loops play in complex systems?
- Why might connectivity be important for life’s emergence?
- Could autocatalytic networks evolve before genes existed?
- How might protocells stabilize catalytic networks?
- What limitations arise when modeling networks using simple abstractions?
- Can complexity emerge without selection?
- Are autocatalytic networks sufficient for life, or merely one component of it?
- Could life’s origin have required both networks and replicators simultaneously?